
December 31st. 2009
We now know that DNA has at least three levels of indirection in its operation. At the lowest level are genes that directly encode proteins; when these genes are activated, the DNA in that gene unzips, builds a protein, ejects it, and re-zips. The protein meanders out into the cell body where it reacts with other proteins, making wonderful things happen.
At the next level are "regulator genes", each of which lies close to a base-level gene. That regulator gene controls the base-level gene, activating or de-activating it under appropriate conditions.
At the third level are regulator genes that I will call "lieutenant regulator genes"; they activate groups of regulator genes. These lieutenant regulator genes apparently express phenotypes by turning on or off entire groups of genes that, operating as a group, handle some bigger task. Above them are "captain regulator genes", "major regulator genes", "colonel regulator genes" and so forth For example, let’s consider an entirely imaginary general regulator gene that builds the little toe of a embryo. When activated, it turns on the group of colonel regulator genes that activate the major regulator genes that activate the captain regulator genes that activate the lieutenant regulator genes that activate the individual proteins required for making a toe. Of course, you don’t make a toe out of a single cell; instead, what happens is that cells at the proper location for the toe should activate their "little toe general regulator gene" so that they build a little toe. Cells anywhere else in the embryo do not activate this gene.
Here is the insight I had (which was probably figured out by embryologists long ago): This is all part of a huge tree of embryological development. At the very top is an "Commander-in-Chief Regulator Gene" that, in effect, codes the commands for "begin construction of a body". It activates at conception, activating genes for growth and cell division, and sets up the chemical gradient that embryos use to determine where the top and bottom of the body lie. Those genes in turn activate lower-level genes (the HOX genes) that lay out the basic structure of the body: head, spine, and limbs in the typical vertebrate. An even lower level of HOX genes then specifies medium-level details of body structure by activating an even lower level of HOX genes... and so on throughout development. Thus, the logical structure of DNA operation looks something like this:
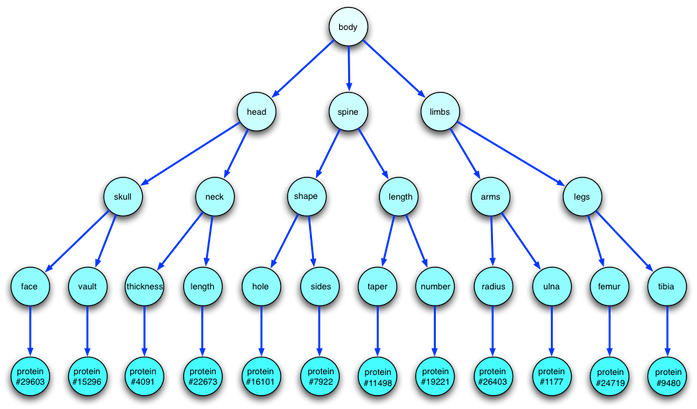
I emphasize that this is a purely mental construct; it has no physical correlate. The structure of this tree does not in any way correlate to the structure of DNA, nor does it correlate with the "tree of life". It is a time-ordered structure, showing how the "program" for building bodies works. In this sense, it’s rather like a flow chart for a computer program.
However, there is one aspect of this tree diagram that does correlate with something real. Let us bring to bear the half-correct dictum that "Ontogeny recapitulates phylogeny". (I love that sentence because it is perhaps the most information-dense statement I have ever encountered.) This diagram is an ontological flow chart -- it shows the logical process by which an fertilized egg becomes a newborn creature. So, is there some sort of phylogenic structure here as well? I believe that, in a rather loose sense, there is just such a phylogenic structure. The earliest, simplest creatures used a tree structure like this, but it was a subset of this structure. Evolutionary processes altered this tree in three ways: broadening it, deepening it, and altering its internal details. Remember that every step had to be done with just an incremental change in DNA.
Every gene in DNA builds a particular protein. Base-level genes produce proteins that travel out into the cell and make wonderful thing happen. Control genes, at any supervisory level, operating by making proteins that activate or de-activate other genes. Random mutations alter random bits of DNA, causing the gene that owns that bit to produce an incorrect protein. In some cases, this has no discernible effect; the change in the protein does not affect its operation. In most cases, however, that protein is altered in a manner that prevents it from performing its function; cells with such mutations in them will likely die or malfunction. Either way, it’s bad for the creature. But in some rare cases, the altered protein just happens to activate (or deactivate) another gene, and that gene activates or deactivates a whole cascade of changes.
Let me coin some clarifying terminology: subtrees. A subtree is a subset of the big tree, but it’s a proper tree of its own. It has a single commander gene at the very top, which regulates some genes below it, which in turn regulate even lower genes. You could think of subtrees as akin to military units: squads, platoons, companies, battalions, and so forth. And at the bottom of every subtree are the lowly privates who make the proteins that go out into the cell and make wonderful things happen.
Here’s an example of my meaning. Suppose that by some wild accident, the gene that manufactures protein #4092 gets clobbered by a hydroxyl radical that knocks out a crucial link, so that the gene now produces a completely new protein, #39999. Suppose further that, by some equally wild happenstance, protein #39999 happens to be the activation protein for the gene at the top of the subtree for making legs. That starts the process of growing a new leg. Nine months later the National Enquirer headlines a story about a three-legged baby.
Three-legged humans probably can’t walk, so that mutation would have died out immediately. But many other mutations that accidently activate regulator genes at any command level are certainly possible. Note that they cannot create completely new trees out of thin air; they can only activate subtrees in different ways. Perhaps they temporarily deactivate the subtree for making thigh-bones, but later the mutated gene is deactivated by a higher level gene. Without the deactivation protein, the thighbone starts growing -- but now it has less time to grow and so it ends up being smaller than a normal thighbone. Similarly, if the mutation causes the thighbone gene to start to work sooner, then the thighbone will be longer.
Now I must confess to you that I lied about making three-legged babies. That’s almost impossible, because genes are not switched on or off by single regulators. In the first case, a cell must be pluripotent (pluripotence is what makes stem cells so special; but stem cells differ from each other in the degree and nature of their pluripotence.) An adult of any species has very few stem cells, so if a cell in my nose gets zapped by a gamma ray that mutates it in such a way as to activate the "make a nose" subtree, I won’t end up with two noses; my nose cells aren’t pluripotent, so they can’t grow into the various cells required to make a nose.
Nevertheless, the concepts I used in that horrid example are sound. Evolution takes place by the random generation of new proteins that have new effects upon the operation of the cells in the body.
It is also possible to extend the tree downward; this is how DNA grew from the short strands that we assume existed in early forms of life into the very long strands that modern creatures possess. For this to happen, a gene must be mutated in such a way as to create a completely new protein that changes the operation of the cell as a whole. Let me offer another purely speculative example. Suppose that we have some genes for making hair. We want hair to be a strong strand of proteins, capable of handling plenty of wear and tear without breaking or falling off. Suppose that our exemplar creature is making pretty good hair with a "private" gene that makes a pretty good protein. But now suppose that a completely different private gene somewhere in the chromosomes is mutated in such a way that it starts manufacturing a protein that just happens to trigger a previously dormant section of DNA; that newly-active stretch of DNA just happens to make a protein that’s better for making hair with. This is complicated, so let me trace it out again: the cell already has a perfectly good private (as opposed to lieutenant, captain, etc) gene that makes decent hair. But now suppose that a previously dormant section of DNA (there’s a lot of apparently dormant DNA in most genomes; scientists think that it functions as raw material for mutations such as this) gets zapped in such a way that it produces a superior protein for making hair, and also just happens to be activated by one of the proteins manufactured by a private-level gene. Sure, this requires a coincidence between the "superior hair protein" feature and the "activated by an existing protein" feature, but we’ve had zillions and zillions of trials, and it obviously took several million years for mammals to get really good hair. So while it’s improbably singly, it was highly likely in the long run.
Here’s the key point of this example: a gene that was previously a private now discovers that its protein is a regulator protein. The private just got promoted! Or, to put it another way, the tree was extended downward by one step. I confess that this is a pretty lame example, but I think it sufficient to make the point that these trees should be able to grow downward as well as sideways.
To conclude: early cells started off with short DNA, but over millions of years that DNA lengthened as the operational tree for its functioning grow larger. It was a slow, incremental process, adding one new leaf to the tree every now and then, branching that leaf off to start a new subtree, and so forth.